Lee HS, Lee BC, Kang DI. Spontaneous self-assembly of DNA fragments into nucleus-like structures from yolk granules of fertilized chicken eggs: Antoine Béchamp meets Bong Han Kim via Olga Lepeshinskaya. Micron. 2013 Aug;51:54-9. doi: 10.1016/j.micron.2013.06.009. Epub 2013 Jul 5. PMID: 23916568.
「鶏受精卵の卵黄顆粒から核様構造へ自発的自己集合するDNA断片~アントワーヌ・べシャンは、オリガ・レペシンツカヤを介してキム・ボンハンと邂逅する」
要旨
卵黄顆粒からのDNA分子の自発的な自己集合が、卵の受精のごく初期段階で起こったという証拠を発見した。DNA分子の自己集合の確かな証拠を見つけるために、受精卵のさまざまな段階で入手可能な多くのデータを収集し、データ表を作成した。まず、アクリジンオレンジのバイタル染色を用いてDNAを証明したところ、いくつかの卵黄顆粒がDNAシグナルを発しており、そのシグナルはインキュベーション時間が長くなるにつれて、非常に小さなサイズから非常に大きな核のような構造へと徐々に増加していることに気づいた。より確実な証拠とするため、別のバイタル色素であるHoechst33258 DNA特異的色素も用いて、卵黄顆粒の変化を追跡した。Hoechst 33258で染色した卵黄顆粒からのDNAシグナルのパターンは、アクリジンオレンジで染色した卵黄顆粒からのものと同じであった。卵黄顆粒の変化を部分的な位相差顕微鏡像で観察したところ、核様構造が形成される前に、顆粒の周囲に液体のようなものが見られた。蛍光顕微鏡と部分位相差顕微鏡を併用することで、これらの液体状の物質は、DNA分子の自発的な自己集合が起こった卵黄顆粒から放出された可能性が示唆された。最後に、DNAシグナルが本物のDNA分子によるものかどうかを検証するため、デオキシリボヌクレアーゼI(DNAse)を用いて、核様構造物が本当にDNA分子が集合したものであることを確認した。この論文では、DNA分子が細胞のような構造に向かって自己集合していることを示す証拠を報告するとともに、われわれの発見を、他の先駆者たち、特に新しい細胞を生み出すための有糸分裂を伴わない代替経路の存在を主張したアントワーヌ・ベシャン、オルガ・レペシンスカヤ、キム・ボンハンの研究成果と比較しながら論じる。
導入
生物学の歴史には異端と見なされた科学者が数人存在する。アントワーヌ・べシャン(Bechamp, 1912)、ガストン・ネサン(Davies, 1991)、ウィルヘルム・ライヒ(Wilehlm, 1979)、オルガ・レペシンツカヤ(Lepeshinskaya, 1954)、キム・ボンハン(Kim, 1965a, Kim, 1965b)などである。彼らは共通して、生命の最小単位は細胞ではなく微粒子だと主張したことが無視されている。その微粒子に各々独自に命名している:マイクロザイマ、バイオン、ソマチッド、生体物質、サンアルズ。しかし、彼らのアイデアは、微粒子を含有するDNAの集合を介した、細胞の有糸分裂を伴わない細胞新生の代替経路であるという点で共通している。彼らの内、我々の関心は2002年から現在に至るまでのキム・ボンハンの研究に焦点が当てられた。まず我々は、キム・ボンハンが主張したように、鍼灸経絡系に対応する解剖学的構造:血管に浮遊するプリモ血管(PV)とプリモ結節(PN)(Lee et al., 2004, Lee et al., 2008)、ウシの心臓(Lee et al., 2011)、リンパ管(Johng et al., 2007, Lee et al., 2006)、脳室と脳静脈洞(Lee et al., 2010, Lee et al., 2012a)の実現を調査した。ごく最近、著者の一人であるBC Leeは、PVとPNからなるシステムであるプリモ血管系(PVS)が微粒子を含有するDNAを含み、輸送する可能性があることに気づいた(Lee et al., 2013a)。
我々のこれまでの研究に基づいて、融合したDNA構造体が細胞としての役割を果たすことを証明することはできなかったが、微粒子を含有するDNAの集合と融合を証明した(Lee et al., 2012b, Lee et al., 2012c, Lee et al., 2013b)。興味深いことに、最近、DNAナノ粒子の3D構造体への集合を報告した文献が数件ある。しかし、その集合は人工的な条件下であって、生体内での現象ではない(Ke et al., 2012, Sudhof, 1995)。この時点で、生体のDNA断片が、特定の条件下で集合するのではないかと推測した。参考文献の研究から、卵黄顆粒自体にゲノムDNA断片が含まれていることに気づいたが、その機能は謎のままだった。さらに、先駆者の一人であるオルガ・レペシンスカヤは、卵黄顆粒を新しい細胞を作るための生体物質と考えた。これまでの研究成果や参考文献から、我々は卵黄顆粒中のDNA断片が核の集合に重要な役割を果たしているという作業仮説を提唱した。
我々の作業仮説を調査するべく、先行研究同様、DNA断片のインキュベーションと観察が同時に実施可能な温度制御システムを備えた顕微鏡を挿入した特殊なチャンバーを作製した。検体は、ホモジナイズした胚盤葉と卵黄顆粒が多数集まる受精卵の暗域(area opaca)から採取した。検体のDNAシグナルを追跡する為、アクリジンオレンジとHoechst33258の2種類のDNA生体染色色素を用いた。また、また、部分位相差顕微鏡を使って、位相差画像だけでなく、卵黄顆粒の明視野画像も観察した。最後に、そのDNAシグナルが、実際のDNA分子に由来するか確認する為にデオキシリボヌクレアーゼ I(DNAse)を用いた。一連の実験により、インキュベーション時間の経過中におけるDNAシグナルの詳細な画像変化を追跡することが可能となった。以上より、我々は、卵黄顆粒から放出されたであろうDNAが集合する可能性を示唆する。また、最初の先駆者であるオルガ・レペシンツカヤの研究と我々の卵黄顆粒の研究との類似性と差分についても議論する。
素材と手法
2.1. 観察の準備
これらの実験のために、検体の観察とインキュベーションに特殊な準備を実施した。詳細は先行研究に記述している (Lee et al., 2013b)。
2.2. ニワトリ受精卵からの検体作製
本研究のために、新鮮な受精卵(それぞれ約60g)Nong Hyunp社(Deajeon, Korea)から取得した。卵を自動デジタルインキュベータ(Boohoacheonguk, Deajeon, Korea)で培養し、これにより卵を37.2℃~38.0℃の比較的一定温度に保温した。正確な検体を特定するために、受精卵から胚盤葉または暗域を様々な段階で分離した。また、ゴム棒を使って手作業でホモジナイズした。検体作製はコンタミネーションを避けるため、すべてクリーンベンチで行った。
2.3. 培養中の検体の観察
リン酸緩衝生理食塩水(PBS, pH = 7.4)に0.01%のアクリジンオレンジ若しくはHoechst 33258を1-2滴添加した。その後、図1に図示するように、スライドガラス上のワセリン壁に約20μlの試料を装填した。準備した検体を装填する前に、先行研究(Lee et al., 2013b)で示したインキュベーション・システムが最適温度(約37.5℃)で作動していることを確認した。その後、(CCD)を使用し、卵黄顆粒の変化を数時間観察した。光学的アーティファクトを除去するべく、全ての写真を同じISO1600で撮影し、培養時間中の卵黄顆粒の変化を比較した。最後に、DNAse(1 mg/ml, デオキシボヌクレアーゼ I, ウシ膵臓由来、比活性2200 Kunitz units/mg, SIGMA)の効果を確認するため、受精卵の暗域から採取した同じ検体を、対照検体(DNAseなし)と試験検体(DNAseあり)に分けた。このセットアップでは、核様シグナルが本当に集合したDNA分子に由来するか確認するため、DNAseの最大使用濃度である1mg/mlを使用した。2時間のインキュベーション後、対照検体と試験検体の核様構造の変化をHoechst 33258染色を用いて調べた。
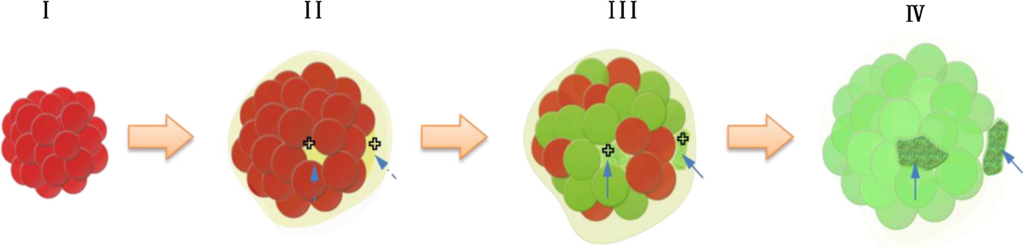
・第一相は卵黄顆粒のクラスターを示しており、約1時間のインキュベーションの後、卵黄顆粒は液体状の物質を放出する。
・第二相で示すように、放出された液体状物質(淡い黄色)は、点線の矢印で示すように2つのぼやけたクラスターがあり、その一部はほとんど視認できない。
・第三相は液体状物質の内部にある2つのクラスターを示しており、緑がかったように見られ、DNAが卵黄顆粒の一部で色を変化させていることが示唆される。
・第四相で示すように、数時間のインキュベーションの後、2つの矢印で示す特徴的なDNAシグナル含有構造が出現する。同時に、全ての卵黄顆粒が完全に赤色蛍光から緑色に変化する。特に、DNAリガーゼ(十字バー)は、第二相と第三相でDNAの集合のために活発に作用していると推測される(この図の凡例中の色に関する言及の解釈について、読者はウェブ版の論文を参照されたい)。
3. 結果
先行研究(Lee et al., 2013b)で使用した特殊なセットアップを用いて、受精卵から採取した卵黄顆粒の培養と変化を観察することができた。予備データとともに、まず図1に示したような作業仮説を立てた。図1に示す通り、卵黄顆粒はDNA断片を持つことから、DNA物質を放出すると推測した。
図2は、アクリジンオレンジで染色した受精卵の暗域から採取した卵黄顆粒クラスターのDNAシグナルが、23時間以上かけて徐々に変化する蛍光画像である。インキュベーションの前には、全ての卵黄顆粒クラスターでDNAシグナルは全く検出されないか、微量にしか検出されていない。インキュベーション後、DNAシグナルの強度が、クラスターの直径の増大とともに徐々に増加した。画像(A)はDNAシグナルが一切出現していないことを示しており、1,2,23時間のインキュベーションでシグナルが徐々に増幅していた。また、卵黄顆粒の色も赤色蛍光から緑色に変化した。別の代表的な変化は、最初はDNAシグナルが無かった2つの卵黄顆粒クラスターで、インキュベーション時間の経過とともにDNAシグナルが徐々に出現し、強くなったことである。
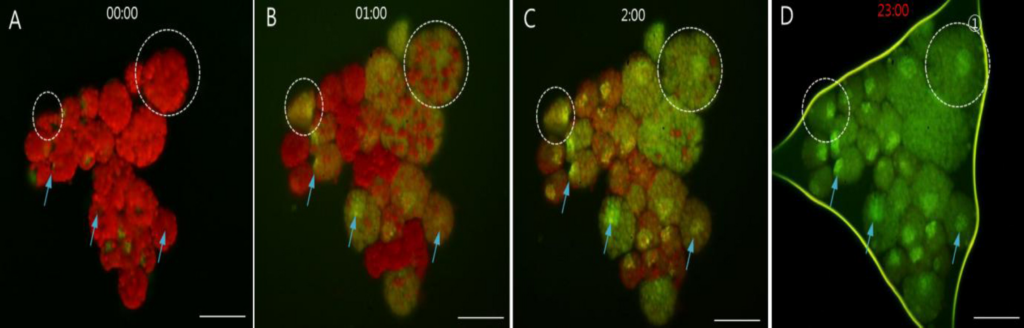
(この図の凡例中の色に関する言及の解釈については、読者はウェブ版の論文を参照されたい)。
DNAシグナルの変化を特異的に追跡するため、図3に示したようにHoechst 33258を使用した。Hoechst 33258で染色した検体のDNAシグナルの変化のパターンは、図4のアクリルオレンジ染色の検体と同様であった。画像(A)はインキュベーション前の卵黄顆粒クラスターの画像であり、僅かなDNAシグナルが一ヵ所観測された。一時間のインキュベーションの後、別の僅かなDNAシグナルが出現したが、他にも視認困難なDNAシグナルが存在したものと思われる。更に1時間の後、観測されたほぼ全てのシグナルが、明るいDNAシグナルを伴う特徴的な構造を示した。
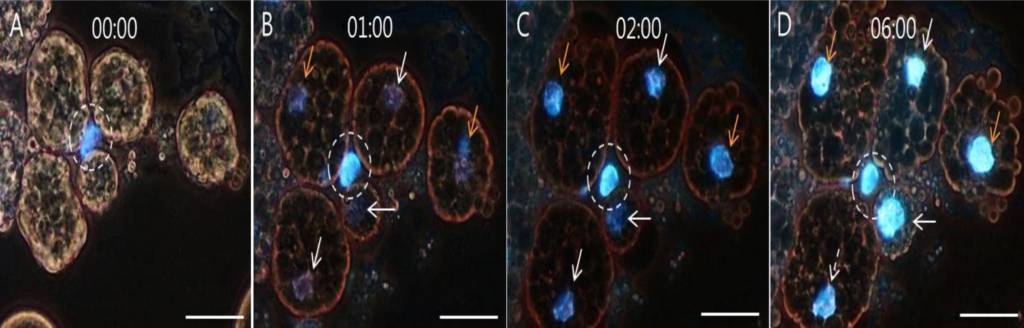
(この図の凡例中の色に関する言及の解釈については、ウェブ版の論文を参照されたい)
図4は23時間以上に亘るインキュベーションで、アクリジンオレンジで染色した卵黄顆粒クラスターのDNAシグナルが徐々に変化していく様子の部分位相差と蛍光染色画像である。卵黄顆粒クラスターの部分位相差画像は、卵黄顆粒クラスター周辺に液体状物質の出現を示唆している。特に、液体状物質は培養時間の経過とともに透明から黄色に変化し、新たな液体状物質も出現した。部分位相差顕微鏡下の液体状物質は、緑色蛍光顕微鏡下の液体状物質に対応する。蛍光顕微鏡データは、徐々に強まるDNAシグナルの存在を証明した。
(1)DNAシグナルがない状態から、卵黄顆粒からの微弱なDNAシグナルを経て、強力に集合したDNAシグナルに変化
(2)卵黄顆粒の外側の液体状物質から、いくつかの微弱なDNA緑色シグナルが出現し、1時間の培養期間中に徐々に強まっていった。最終t系に、14時間の培養後、特定の構造の中でDNAシグナルが更に強化された。
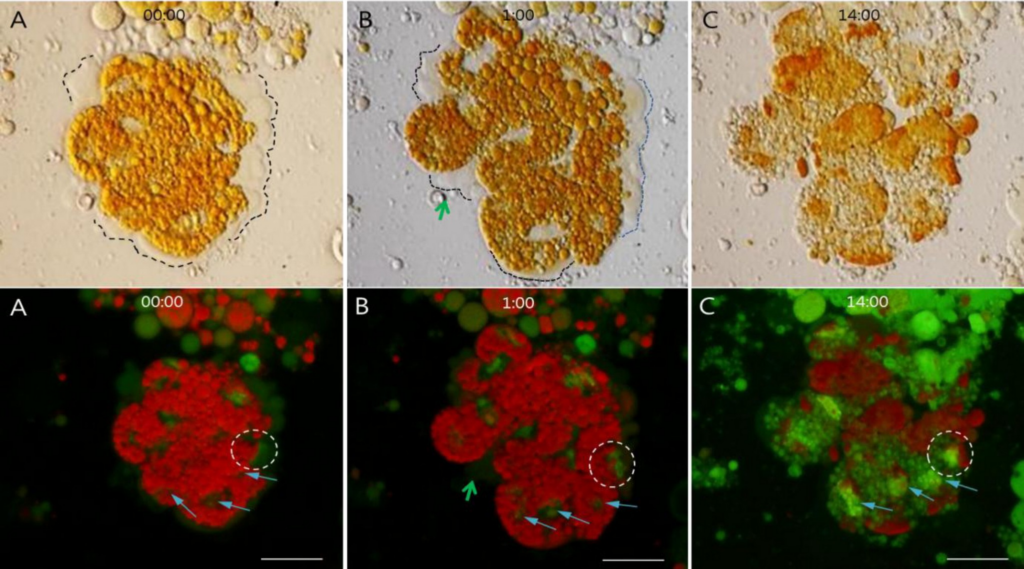
卵黄顆粒クラスターの部分位相差画像は、クラスター周辺の点線で示される液体状物質の存在を示唆している。特に、培養期間を経て液体状物質の色が透明から黄色に変化し、(B)の緑色矢印で示される新たな液体状物質も出現した。部分位相差顕微鏡下のこれら液体状物質は、下段の緑色矢印が示す緑色蛍光に対応する。下段はDNAシグナルが徐々に強化する様子を表している。
(1)3つの細い矢印は、(A)DNAシグナルが無い状態から、卵黄顆粒からの微弱なDNAシグナルを経て、(C)強力に集合するDNAシグナルへの変化を示している。
(2)点線の円で示される卵黄顆粒の外側の液体状物質に微弱なDNAシグナルが存在し、1時間以上の培養を経て(B)徐々に強まっていった。最後に、14時間の培養を経て、(C)の点線の円にあるように、DNAシグナルが強力になり、濃縮した構造となった。スケールバーは全て25μmである。
(この図の凡例中の色に関する言及の解釈については、読者はウェブ版の論文を参照されたい)。
図5はHoechst 33258で染色した卵黄顆粒クラスターのDNAシグナルが4時間の培養で徐々に変化する様子を、部分的な位相差画像と蛍光画像で示したものである。卵黄顆粒クラスターの位相差画像(A)は、クラスター周囲に液体状物質が存在することを示唆している。蛍光画像は、4時間の培養でDNAシグナルが徐々に強まったことを示している。
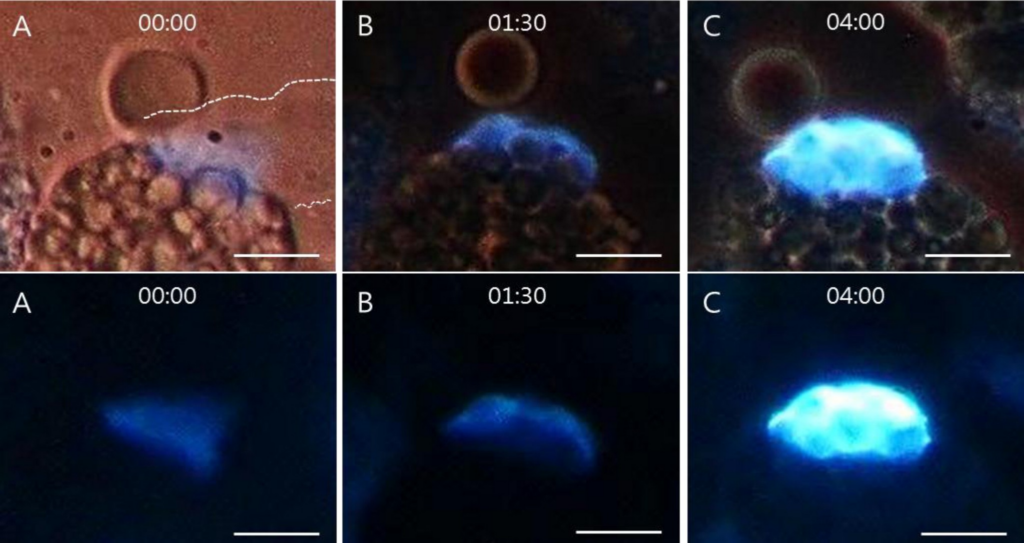
核様構造が本物のDNA分子であることを確認するため、対照として同じ検体にDNAseを適用した後の核様構造の変化を調べた。図6に示すように、Hoechst 33258で染色した卵黄顆粒クラスター周辺の核様構造を2時間インキュベートした後、位相差画像と蛍光画像を撮像した。対照検体の位相差画像A及びBと、試験検体C(DNAse処理)の位相差画像は同じ特徴を共有していた。しかし、蛍光画像は決定的に異なっており、対照検体A及びBでは特徴的なDNAシグナルが検出された一方、DNAse処理検体のCではクラスターからの蛍光は見られなかった。DNAseの影響を確認するため、DNAの蛍光を示さなかったスライド上の全ての検体を観察した(図6のCを代表例として示す)。
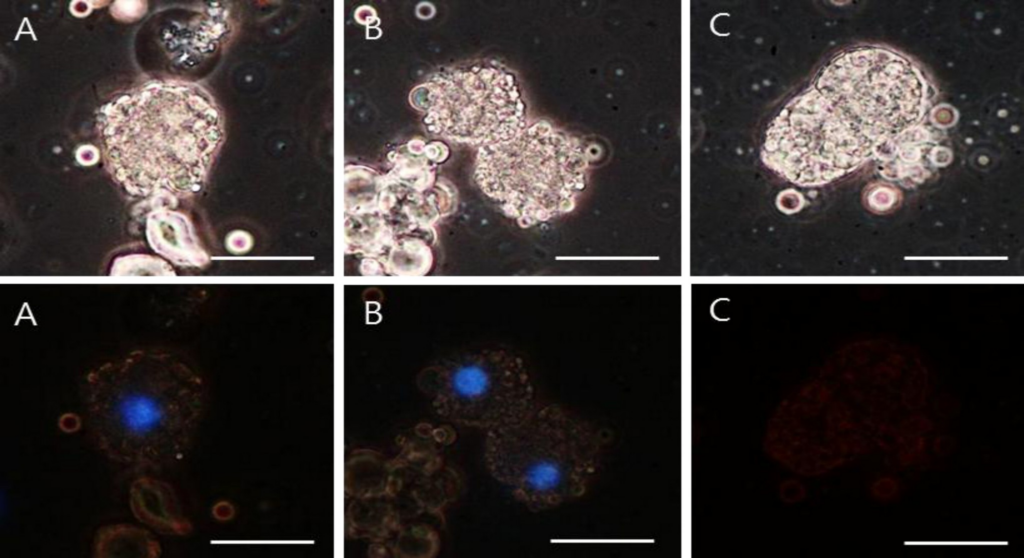
位相差画像(上段)A,BはDNAse未処理の対照検体だが、画像CはDNAseで2時間インキュベート処理した検体の試験画像である。全ての位相差画像は同じパターンを共有している。
蛍光画像(下段)は異なる画像である:対照検体A,Bは特徴的なDNAシグナルを示しているが、DNAse処理検体Cは蛍光を示さず、DNA分子が形成されなかったことを示唆している。スケールバーは全て25μmである。
4.考察
これらの結果を総括すると、卵黄顆粒クラスターは、DNA分子の自発的自己集合を経て、卵の受精のごく初期段階において、新たな核を生成する上で極めて重要な役割を担っている可能性が示唆された。生物学的に、これら卵黄顆粒クラスターは”周縁質(periblasts)”と命名されているが、核との機能的な関連は未解決のままであり(Yamamoto, 1982)、従って、本報告ではこれらを鶏受精卵中の卵黄顆粒のクラスター形態をみなした。
表1に示す通りの一連の実験を通して、有糸分裂とは異なる細胞新生の代替経路が存在する可能性があるという暫定的な結論に達した。
(1)卵黄顆粒から液体状物質が放出され、その中でDNAの自発的自己集合が生じ、核様構造が形成される。
(2)卵黄顆粒からのDNA分子が集合すると、卵黄顆粒の色が赤色蛍光から緑色へ変化し、これは卵黄顆粒に関する以前の研究(Fausto et al., 2001)と同様の意味合いを持つ。
(3)DNAse処理により、核様構造が、実際にDNA分子が集合が集合したものだと確認された。
表1.
| 番 | 培養時間(h) | 観察時間(h) | 検体 | 染色 | 液体状物質の有無 |
| 1 | 0 | 23 | 胚盤葉 | アクリルオレンジ(AO) | × |
| 2 | 13 | 57 | 胚盤葉 | AO | 〇 |
| 3 | 6 | 14 | 胚盤葉 | AO | 〇 |
| 4 | 21 | 18 | 胚盤葉 | AO | 〇 |
| 5 | 28 | 4 | 胚盤葉 | AO | 〇 |
| 28 | 4 | 胚盤葉 | AO | 〇 | |
| 6 | 0 | 1.5 | 胚盤葉 | AO/Hoechst 33258(H) | × |
| 7 | 0 | 2 | 暗域 | AO/H | × |
| 8 | 0 | 1 | 暗域 | AO/H | × |
| 9 | 0 | 6 | 暗域 | AO/H | × |
| 0 | 4 | 暗域 | AO/H | × | |
| 10 | 0 | 4 | 胚盤葉 | H | 〇 |
培養時間とは、受精卵を培養してから検体を採取するまでの時間である。
観察時間は卵黄顆粒の変化を追跡した時間を示す。
色素中のAOはアクリジンオレンジ、HはHoechst 33258、AO/HはアクリジンオレンジとHoechst 33258の組み合わせを示す。
LLMは卵黄顆粒から放出される液体状物質を意味する。
卵の受精のごく初期にDNA断片が自発的に核に集合するという我々の発見は全く新しいものだろうか?我々の発見以前、卵黄顆粒の重要性を最初に認識した先駆者はオルガ・レペシンスカヤであった(Lepeshinskaya, 1954)。彼女は、我々の推測と同じく卵黄顆粒が新たな細胞を生成する生体物質だと主張したが、彼女は卵黄顆粒自体が新細胞に変化すると考えた。これとは対照的に我々は、ゲノムDNA断片を持つ卵黄顆粒が、DNA断片を集合させて核を形成するための供給源として機能すると考えている。また、彼女は、卵黄顆粒が液体状物質を含有するDNA物質を放出することに気づかなかったが、我々は部分位相差顕微鏡下で液体状物質の出現を確認した。更に、我々は液体状物質の内部から新たなDNAシグナルの形成を観察した。この時点で、我々が主張したように、卵黄顆粒以外の種類の細胞からもDNA分子が放出されるかと考える人がいるかもしれない。興味深いことに、生命科学者たちは、リンパ球やがん細胞、さらにはバクテリアからもDNA分子が放出されることに気づいている(Chen et al., 2005, Skvortsova et al., 2006, Pisetsky, 2012)が、細胞外DNA(eDNA)と命名されている、放出されたDNA分子の機能は謎のままである。以上より、我々は、卵黄顆粒から放出された液体状物質と細胞から放出されたeDNAが同じ生物学的機能を有していると示唆する。一方、卵黄顆粒のゲノムDNAのDNAシーケンスは不均一であることが報告されている(Lijun et al., 1998). 従って、理論的には、異なる卵黄顆粒が異なるDNA断片を放出し、これが集合して細胞内の核に必要なDNA情報の全てを画乳する核様構造体を形成する可能性がある。しかし、核様構造が、実際の核や細胞の役割を果たすかどうかは確認できていない。興味深いことに、鍼の経路の解剖学的実態を明らかにした先駆者であるキム・ボンハンも、新たな細胞を生成する最初の事象は核様構造の形成であり、そこから細胞質が自然に形成されると主張していた。一方、現代科学のホットイッシューの一つである合成生物学は、様々な人工的アプローチによりDNA集合に依存している(Ke et al., 2012, Sudhof, 1995);。皮肉にも、DNA断片の自発的集合が生体内で生じることを複数の先駆者がすでに予見していた (Bechamp, 1912, Davies, 1991, Wilehlm, 1979, Kim, 1965b)。
この時点で、DNAの自発的自己集合は受精卵のごく初期にしか発生しないのだろうかと思うかもしれない。我々の先行研究では、DNA断片が同心円状に成長し、より大きなDNA断片となることを示唆している(Lee et al., 2012c)。ごく最近、我々は、マイクロベシクル内のDNA断片が自発的に結合し、正常細胞と同じ形態を持つ特定構造になるという証拠を示した(Lee et al., 2013b)。関連の研究で、DNAシグナルを含有するマイクロベシクルが、異なる種の様々な組織から採取できることに気づいた(Lee et al., 2013b)。我々の先行研究と先駆者の研究に基づき、我々はDNA断片の自発的自己集合があらゆる生物種で生じうると考えている。この新たな現象を理解するためには、ワトソンとクリック(Watson and Crick, 1953)が示唆した半保存的複製以外の新たな遺伝理論の構築が必要であることを示唆している。
この新たな現象を臨床応用するためのより重要な研究のために、1880年代のアントワーヌ・べしゃん、1950年代のオリガ・レペシンスカヤ、1960年代のキム・ボンハンの3人の先駆者の洞察を改めて紹介したい。アントワーヌ・ベシャンは、マイクロザイマと命名した微小粒子が集合して新たな細胞を生み出すと示唆した最初の先駆者である(Bechamp, 1912, Lepeshinskaya, 1954, Kim, 1965b)。オリガ・レペシンスカヤは、これら微小粒子が新しい細胞へ集合しうると主張し(Lepeshinskaya, 1954)、キム・ボンハンは、サンアルズと命名したDNA断片が流れる鍼の経絡系に対応する新しいシステムを示唆した(Kim, 1965b)。以上より、DNAの自発的自己集合の証拠に関する我々の発見は、鍼の経絡系と推定されるプリモ血管系という形で、新たな遺伝的原則を持つ普遍的パラダイムの到来を告げる可能性を示唆している。アントワーヌ・ベシャンは、オリガ・レペシンスカヤを介してキム・ボンハンと邂逅する。
謝辞
本研究は、2010年に韓国教育科学技術部(MEST)より支給された韓国国立研究財団(NRF)助成金(No.2010-0025289)および韓国薬局方研究所より支給された韓国薬局方財団助成金の支援を受けた。また、Jeong Yim Lee女史の技術協力に感謝する。